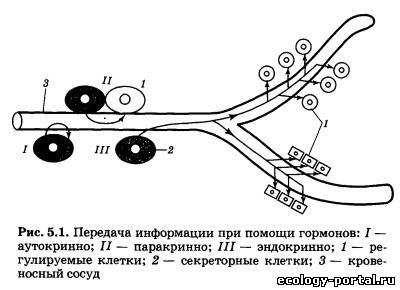
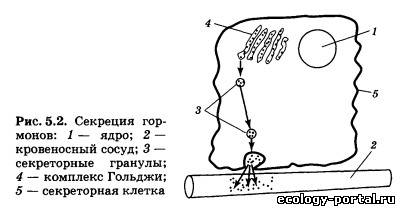
Особенностью многоклеточных животных является диф- ференцировка тканей, причем каждый вид тканей приспособ- лен к выполнению специализированных функций, что обеспе- чивает выживание организма и его адаптацию к непрерывно меняющимся условиям существования. Для того чтобы из миллиардов клеток создать целостный, интегрированный организм, способный к самовосстановле- нию, самовоспроизведению и адаптации, необходима постоян- но действующая система коммуникации между клетками. В ходе эволюционного процесса сформировались две такие системы: нервная и эндокринная. Нервная система, о которой подробно рассказывалось выше, структурирована, и аксоны нервных клеток образуют синаптические окончания в строго определенных местах организма. Эндокринная система ис- пользует для передачи сигналов от одних клеток к другим осо- бые химические вещества — гормоны (от греч. hormao — про- буждать, активизировать). Гормоны — это специальные регу- лирующие вещества, вырабатываемые в одних тканях организма, транспортируемые с током крови к различным ор- ганам и воздействующие на их работу. Гормоны, несмотря на свое название, далеко не всегда «возбуждают» регулируемый ими орган, и характер их воздействия может очень сильно варьировать. Как известно, нервные импульсы распространя- ются по аксонам с большой скоростью, и для осуществления, например, коленного рефлекса необходимы доли секунды. В противоположность этому гормональная регуляция осу- ществляется гораздо медленнее, и под ее контролем находятся те процессы нашего организма, которые требуют для регуля- ции минуты или часы. Передача информации при помощи гор- монов может осуществляться в пределах организма на самые различные расстояния. Выделяют три основных вида гормо- нальной регуляции (рис. 5.1): аутокринная I, когда гормон действует на те клетки, в которых он вырабатывается; парак- ринная II, когда гормон действует на близко расположенные клетки той же ткани, и эндокринная III, когда гормон разно- сится с током крови по всему организму и действует на раз- личные его органы. Такое деление эффектов достаточно отно- сительно, и во многих случаях один и тот же гормон может действовать и паракринно, и эндокринно. Гормоны обычно образуются в особых секреторных клет- ках, которые или образуют компактные органы — железы, или расположены по одной или группами внутри органов. Секре- торным клеткам свойственны некоторые морфологические осо- бенности. Обычно синтез и «упаковка» гормонов происходит в одной части клетки, а их выброс в кровь — в другой. Чаще все- го синтезируемые гормоны накапливаются в комплексе Гольд- жи — основном «складском помещении» клетки. Комплекс Гольджи представляет собой стопку плоских мешочков, стенки которых построены из мембран. По мере надобности гормоны упаковываются в маленькие секреторные пузырьки — грану- лы, которые отпочковываются от комплекса Гольджи и пере- двигаются по цитоплазме к наружной мембране клетки, через которую гормон выбрасывается в кровь (рис. 5.2). Некоторые гормоны, например половые, видимо, не упаковываются в гра- нулы и выходят из секретирующей клетки в виде отдельных молекул. Выброс гормона в кровь происходит не постоянно, а только в том случае, когда к секретирующей клетке прихо- дит специальный сигнал, под действием которого пузырьки высвобождают гормон во внеклеточную среду.
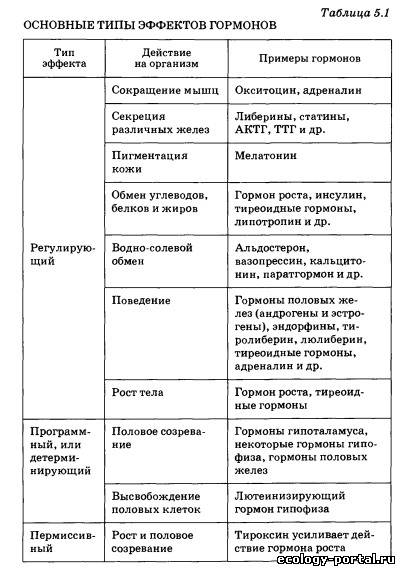
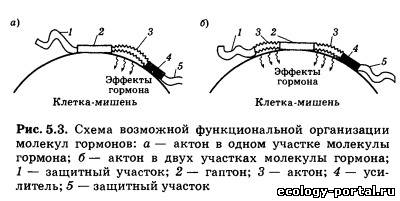
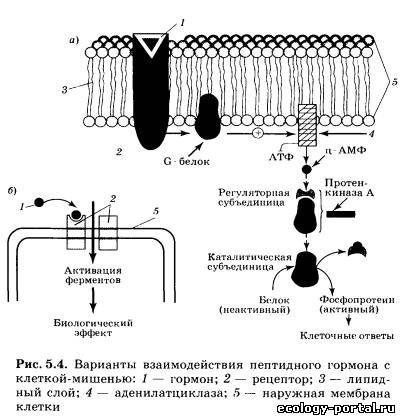
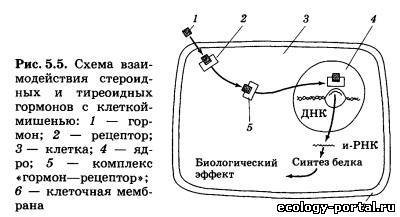
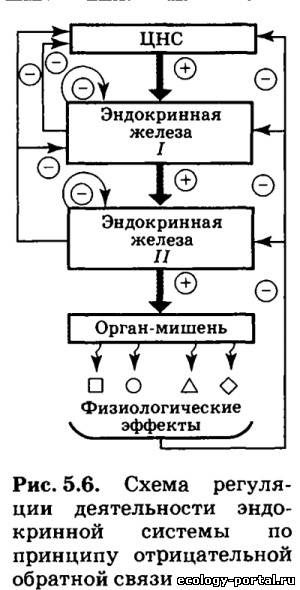
Однако в последние годы стало очевидно, что гормоны смо- гут выделяться не только из клеток специализированных эн- докринных желез, но и из клеток многих других органов и тканей. Так, нейроны гипоталамуса способны вырабатывать целый набор гормональных факторов, таких, как либерины, статины и другие гормоны, клетки сердечной мышцы выделя- ют в кровь натрийдиуретический гормон, лимфоциты выделя- ют ряд гормонов — стимуляторов иммунитета, наконец, мно- жество белковых гормонов синтезируется в слизистой оболоч- ке кишечника. Существует несколько подходов к определению тех крите- риев и свойств, которым должно отвечать вещество, претендую- щее на роль гормона. Перечислим главные из этих критериев. 1. Гормон выделяется из живых клеток. Если какое-то ве- щество, обладающее значительным физиологическим эффек- том, выделяется из мертвой разрушающейся клетки, его к гормонам относить нельзя. 2. Гормоны, как и любые физиологически активные веще- ства, обладают сильными эффектами и поэтому выделяются в очень малых количествах. 3. Гормон выделяется из клетки, не нарушая ее жизнеде- ятельности и целостности. 4. Гормон поступает в межклеточную жидкость и кровь не- посредственно из секретирующих клеток, и эндокринные же- лезы не имеют выводных протоков, как железы внешней сек- реции. 5. Гормон действует только на определенные органы-ми- шени, имеющие специальные рецепторы для взаимодействия с ним. 6. Гормон не является источником энергии и строитель- ным материалом для нужд организма. Эти критерии, предъявляемые к веществу-гормону, доста- точно строгие, и на самом деле имеется ряд исключений, од- нако перечисленные критерии позволяют четко отличать гор- моны от ферментов, витаминов и других веществ. Регулирующие эффекты — это обратимое активирующее или тормозное действие на системы организма, осуществляю- щиеся постоянно в течение всей жизни. Программные, или де- терминирующие, эффекты необратимы и осуществляются в определенные, относительно короткие периоды жизни, на- пример в период полового созревания у человека. При этом гормон определяет кардинальную перестройку клеток, клет- ки меняют свою форму, функции и остаются такими на все дальнейшее время существования организма. Пермиссивное действие гормонов заключается в том, что одни из них, не влияя на какой-то процесс другого гормона, резко усиливают его эффект. Например, гормон щитовидной железы — тироксин необходим для того, чтобы нормально развивались эффекты гормона роста. Даже из краткой табли- цы видно, что гормоны воздействуют на все функции организ- ма, а с другой стороны, один и тот же гормон может непосред- ственно воздействовать на несколько функций. Так, большая часть гормонов способна прямо или опосредованно влиять на возбудимость и эмоции, изменяя поведение человека. Так как передаваемая гормонами информация закодирова- на в строении их молекул, важно знать основные группы хи- мических веществ, к которым гормоны относятся. Первая группа — белково-пептидные гормоны, представ- ляющие собой цепочки различной длины, состоящие из ами- нокислот. К ним относятся либерины и статины гипоталаму- са, гормоны гипофиза, инсулин, натрийдиуретический гор- мон и некоторые другие. Вторая группа — стероиды, представляющие собой произ- водные холестерина. К ним относятся гормоны коры надпо- чечников, гормоны половых желез. Третья группа — гормоны, производные отдельных амино- кислот. Так, производными аминокислоты тирозина являют- ся адреналин — гормон мозгового слоя надпочечников и гор- моны щитовидной железы — тироксин и трийодтиронин. Гор- мон эпифиза мелатонин синтезируется из аминокислоты триптофана. Четвертая группа — производные арахидоновой кислоты: простагландины и некоторые другие гормоны, о которых бу- дет подробнее сказано ниже. Несмотря на огромное различие в химическом строении, гормоны разных химических групп имеют много общего в ор- ганизации функциональной структуры. Так, в идеальном случае в молекуле гормона можно выде- лить адресный участок, или гаптон, отвечающий за при- крепление всей молекулы к месту специфического гормональ- ного воздействия. Гаптон сам по себе никаким биологическим эффектом не обладает. Другой участок гормональной молеку- лы называется актоном. Именно актон вызывает физиологи- ческие эффекты гормона в клетке-мишени, но сам по себе без гаптона актон не может присоединяться к регулируемой клет- ке. И наконец, в состав гормональной молекулы может вхо- дить целый ряд участков, защищающих эту молекулу от воз- действия разрушающих ферментов, усиливающих или ослаб- ляющих гормональный эффект. Эти участки молекулы получили название акцессорных. Актон, гаптон и акцессор- ные участки могут быть расположены в различных участках гормональной молекулы (рис. 5.3, а), а могут и перекрывать друг друга (рис. 5.3, б). Некоторые гормоны способны воздействовать на работу це- лого ряда клеток различных тканей. Другие — способны не- посредственно воздействовать только на один вид клеток. Это различие определяется тем, что для взаимодействия гормона и клетки-мишени необходимо, чтобы в клетке находились специфические рецепторы, позволяющие считывать инфор- мацию, заложенную в этом гормоне. При взаимодействии гормона с рецептором образуется так называемый гормон-ре- цепторный комплекс, который и воздействует на работу клетки. Таким образом, будет ли клетка взаимодействовать с гор- моном и изменять под его влиянием свою физиологическую активность, определяется тем, имеются ли в этой клетке спе- цифические рецепторы к этому гормону. Если рецепторы есть, клетка будет реагировать на гормон, если таких рецепто- ров нет, клетка не будет взаимодействовать с гормоном, даже если этого гормона в жидкости, омывающей клетку, очень много. Надо отметить, что гормональные рецепторы обладают колоссальной избирательностью по отношению к «своему» гормону, прекрасно различая его среди множества других, да- же очень похожих на него молекул и взаимодействуя именно со «своим» гормоном, несмотря на то что концентрация гор- монов в крови всегда очень и очень мала по сравнению с дру- гими классами веществ. Существуют два принципиально раз- личающихся механизма действия гормонов, в зависимости от того, где образуется гормон-рецепторный комплекс: на по- верхности клеточной мембраны или внутри клетки. Белково-пептидные гормоны (первая группа) состоят из це- почек аминокислот, их молекулы велики и, что очень важно, плохо растворяются в жирах, составляющих основу биологи- ческих мембран и мембран, которые окружают любую клетку: это аденокортикотропный гормон гипофиза, тиреотропный гормон, лютеинизирующий гормон, инсулин. Их действие развивается быстро — за минуты, но продолжается очень не- долго. Обычно гормоны этой группы накапливаются в секре- тирующих клетках и быстро высвобождаются из них в слу- чае необходимости. Проникнуть внутрь клетки через липид- ные слои мембраны такие молекулы не могут, и рецепторы к ним расположены на поверхности клетки. При таком виде рецепции .необходимо решить две задачи: во-первых, сигнал воздействия гормона нужно передать внутрь клетки и, во-вторых, усилить этот сигнал, чтобы достичь значительно- го изменения работы клетки. Разберем один из вариантов та- кого взаимодействия гормонов с клеткой (рис. 5.4, а). Каж- дая молекула гормона связывается с одним рецептором на поверхности клетки-мишени. Пространственная структура рецептора при этом меняется, и он взаимодействует с молеку- лой G-белка, также встроенного в мембрану. G-белок в свою очередь активирует фермент аденилатциклазу, способную индуцировать образование большого числа молекул — мощ- нейшего регулятора внутриклеточных процессов (ц-АМФ), которая может влиять на различные реакции, протекающие в клетке. Как один из наиболее вероятных видов воздейст- вия (рис. 5.4, а), он активирует фермент протеинкиназу А, «снимая» с нее регулирующую субъединицу. Освободившись от этой субъединицы, протеинкиназа А начинает присоеди- нять к определенным белкам клетки высокоэнергетические фосфатные группы, т. е. фосфорилирует их. Одни из таких белков являются ферментами, другие входят в состав стенок каналов, которые пропускают в клетку или выпускают из нее различные ионы. Фосфорилированные белки резко изме- няют свою биологическую активность, и вся работа клетки резко активизируется: мышечная клетка будет сильнее сокра- щаться, секреторная клетка будет вырабатывать больше ве- ществ. При такой схеме передачи гормонального сигнала в клетку сам гормон называют первым посредником, ц-АМФ называют вторым посредником, а протеинкиназу А, которая изменяет работу множества белков клетки, называют внутриклеточ- ным эффектором. По ходу описанного каскада реакций про- исходит мощнейшее усиление сигнала. Так, одна молекула адренокортикотропного гормона (АКТГ), взаимодействуя с ре- цептором менее одной секунды, способна активировать около 1000 молекул аденилатциклазы, которые все вместе способны образовать более 1 млн молекул ц-АМФ и т. д. Таким образом, коэффициент усиления в гормональных каскадах способен до- стигать 107—108. Но естественно, белково-пептидные гормо- ны могут и тормозить работу клетки. В этом случае сигнал от рецептора, с которым связался гормон, доходит до особого ви- да G-белка, который не активирует, а, наоборот, тормозит ак- тивность аденилатциклазы. Некоторые пептидные гормоны, также не проникающие внутрь клетки, имеют другой механизм действия (рис. 5.4, б). Связываясь с рецептором на поверхности клетки-мишени, они открывают в мембране канал, через который в клетку пос- тупают ионы кальция. Кальций — активатор целого ряда вну- триклеточных процессов и других протеинкиназ, таким обра- зом, гормон значительно изменяет деятельность клетки. Принципиально иной механизм действия характерен для многих других гормонов (стероидов, гормонов щитовидной железы). Эти гормоны жирорастворимы и легко проникают внутрь клетки через наружную мембрану. В цитоплазме моле- кулы гормона связываются с молекулой рецептора (рис. 5.5). Комплекс гормон — рецептор проникает внутрь ядра клетки и связывается с участком ДНК, «включая» строго определен- ные гены. Идет образование и-РНК, которая выходит из ядра в цитоплазму, где и начинается синтез определенного белка или белков, изменяющих деятельность клетки. Описываемые эффекты развиваются довольно долго и бывают очень дли- тельными, а зачастую являются программными, изменяя ра- боту клетки на все время ее существования. Именно гормоны с «ядерной рецепцией» ответственны за общую и половую дифференцировку, многие поведенческие изменения. Комп- лекс гормон — рецептор реагирует с генетическим аппаратом обратимо, и после распада этого комплекса рецепторы могут использоваться многократно. Обычно всякая эндокринная железа работает на «уровне покоя», выделяя небольшие количества гормона, но при необ- ходимости секреция может тормозиться или, наоборот, резко усиливаться под действием химических стимулов. Этими хи- мическими стимулами могут быть нейромедиаторы, выделяе- мые из окончаний нервных клеток; гормоны, выделяемые другими эндокринными железами; продукты, образующиеся в результате воздействия гормона. Обычно такие регуляции деятельности эндокринных желез осуществляются по прин- ципу отрицательной обратной связи, т. е. повышение кон- центрации какого-либо гормона в крови или реакция кле- ток-мишеней на этот гормон оказывает угнетающее действие на его синтез и секрецию. Как правило, регуляция по принци- пу обратной связи осуществляется по нескольким цепям — с короткой или длинной петлей (рис. 5.6). В случае обратной связи с короткой петлей продукт деятельности органа-мишени или какой-то другой его физиологический эффект угнетающе действуют непосредственно на эндокринную железу, выделяющую гормон-стимулятор органа-мишени. Обратная связь по длинной петле осуществляется по тому же принципу, но содержит большее число составных элементов, один из которых (или их нечетное число) должен быть «тормозным». В случае двух тормозных звеньев в одной цепи отрицательная связь не срабатывает: два «отрицательных» эффекта взаимно «погасят» друг друга. Регуляция по принципу отрицательной обратной связи позволяет осуществлять очень тонкие и гибкие эндокринные воздействия в физиологических пределах без опасения, что выброс какого-то гормона приведет к мощному, лавинообразному эффекту, который может привести к гибели Нервная и эндокринная системы — основные и интегри- рующие системы, делающие организм единым целым и регу- лирующие его в условиях постоянно меняющейся окружаю- щей среды. Но эти системы нельзя противопоставлять друг другу, кроме того, оказалось, что еще на ранних этапах эво- люции живых организмов возникли неиросекреторные клет- ки, объединяющие в себе свойства нейронов и эндокринных клеток. Эти клетки (как нейроны) способны генерировать и пе- редавать по своим аксонам нервные импульсы, и в то же время в их цитоплазме происходит синтез гормонов, являющихся либо пептидами, либо видоизмененными аминокислотами. У млекопитающих большая часть таких клеток сконцентри- рована в гипоталамусе, где они образуют ряд скоплений — ядра. Через множество вставочных нейронов неиросекретор- ные клетки связаны со всеми сенсорными системами и посто- янно получают афферентные стимулы от интеро- и экстероре- цепторов об изменениях во внутренней среде организма и о влияниях внешней среды. Под действием афферентных сти- мулов неиросекреторные клетки начинают усиленно синтези- ровать гормоны и транспортировать их по аксонам к нервным окончаниям в виде секреторных гранул. Сейчас определена скорость, с которой двигаются по аксо- нам эти гранулы: 2800 мм/сутки. По аксонам гранулы с гор- монами двигаются в особые нейрогемальные области, в кото- рых нейросекрет выходит из окончаний и попадает в крове- носные капилляры, а дальше, как и любой другой гормон, переносится с током крови к органам-мишеням. У человека выделяют две важнейшие нейрогемальные области — нейро- гипофиз (задняя доля гипофиза) и срединное возвышение.