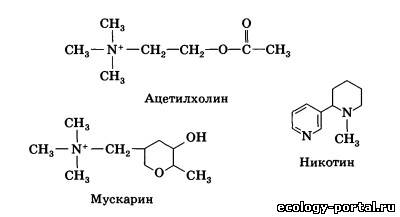
По химическому строению ацетилхолин представляет со- бой соединение двух молекул — азотсодержащего холина и остатка уксусной кислоты:
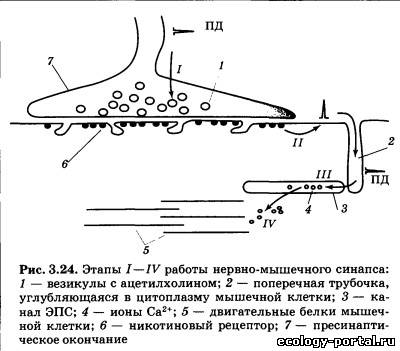
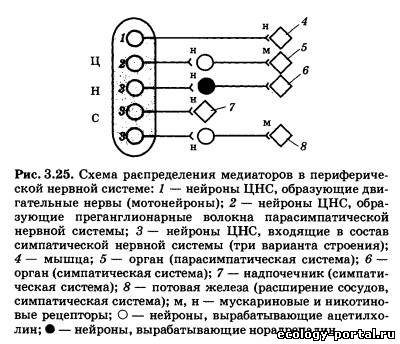
Холин является незаменимым витаминоподобным соеди- нением, получаемым с пищей в достаточном количестве, и по- следствия его дефицита наблюдаются только в искусственных условиях. Синтез ацетилхолина осуществляется в основном в пресинаптических окончаниях с помощью фермента холина- цетилтрансферазы. Затем медиатор переносится в пустые ве- зикулы и хранится в них до момента выброса. Ацетилхолин в качестве медиатора работает в трех функ- циональных блоках нервной системы: в нервно-мышечных синапсах, периферической части вегетативной нервной систе- мы и некоторых областях ЦНС. Ацетилхолин является медиатором мотонейронов нервной системы, которые расположены в передних рогах серого веще- ства спинного мозга и двигательных ядрах черепных нервов. Их аксоны направляются к скелетным мышцам и, разветвля- ясь, образуют нервно-мышечные синапсы. При этом один аксон может устанавливать контакт с сотнями мышечных волокон, но каждое мышечное волокно управляется только одним си- напсом. Размер нервно-мышечных синапсов в десятки раз боль- ше, чем синапсов в ЦНС, и пришедший по аксону мотонейрона даже одиночный ПД вызывает выделение значительного ко- личества ацетилхолина (этап /, рис. 3.24). В результате разви- вающаяся на постсинаптическои мембране деполяризация ока- зывается настолько велика, что всегда запускает ПД мышечной клетки (//), который приводит к выбросу Са2+ из каналов ЭПС (III), активации двигательных белков и сокращению (IV). Периферическое звено вегетативной нервной системы со- стоит из двух нейронов: тело первого (преганглионарного) на- ходится в ЦНС, а аксон направляется к вегетативному ганг- лию; тело второго (постганглионарного) находится в ганглии, а аксон иннервирует гладкие мышечные или железистые клетки внутренних органов. Ацетилхолин в качестве меди- атора вырабатывается во всех преганглионарных клетках, а также в постганглионарных клетках парасимпатической час- ти вегетативной нервной системы (рис. 3.25). Некоторые пост- ганглионарные симпатические волокна (активирующие пото- вые железы и вызывающие расширение сосудов) также секре- тируют ацетилхолин.
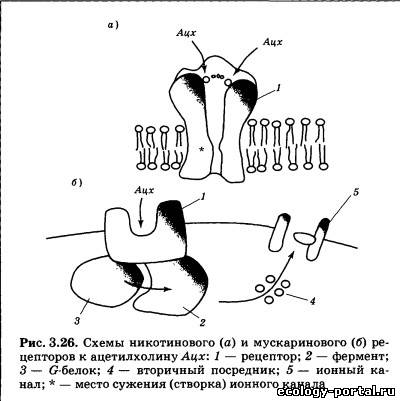
В ЦНС ацетилхолин вырабатывается частью нейронов рети- кулярных ядер моста и интернейронами полосатого тела ба- зальных ганглиев и некоторых других локальных зон. Рассмат- ривается роль этого медиатора в регуляции уровня бодрство- вания, а также в системах памяти, двигательных системах. Доказана эффективность применения антагонистов ацетилхо- лина при ряде двигательных нарушений. Выделяясь из пресинаптического окончания, ацетилхолин действует на постсинаптические рецепторы. Эти рецепторы неоднородны и различаются локализацией и рядом свойств. Выделено два типа рецепторов (рис. 3.26): первый, помимо ацетилхолина, возбуждается под действием алкалоида табака никотина (никотиновые рецепторы), второй тип активируется ацетилхолином и токсином мухомора мускарином (мускари- новые рецепторы). Никотиновые рецепторы являются классическим приме- ром ионотропных рецепторов: их ионный канал входит в со- став рецептора и открывается сразу после присоединения аце- тилхолина. Канал этот характеризуется универсальной про- ницаемостью для положительно заряженных ионов, но в обычных условиях (при открытии на фоне ПП) в связи с нико- тиновыми рецепторами наблюдается в основном входящий Na+-TOK, вызывающий деполяризацию мембраны и возбужде- ние нейрона.
Никотиновые рецептры расположены на постсинаптиче- ской мембране поперечно-полосатых волокон скелетных мышц (нервно-мышечные синапсы); в синапсах вегетативных ганглиев и в меньшем количестве, чем мускариновые рецеп- торы, в ЦНС. Областью, наиболее чувствительной к никоти- ну, являются вегетативные ганглии, поэтому первые попытки курения приводят к значительным нарушениям в деятельнос- ти органов: скачкам артериального давления, тошноте, голо- вокружению. По мере привыкания сохраняется в основном симпатический компонент действия: никотин начинает рабо- тать преимущественно как стимулятор многих систем орга- низма. Присутствует также и центральное активирующее влияние (на головной мозг) ацетилхолина. Сверхдозы никоти- на E0 и более мг) вызывают резкое учащение сердцебиения, судороги и остановку дыхания.
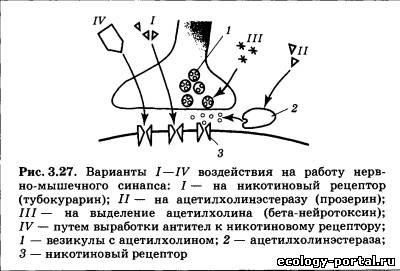
Во время курения никотин действует как слабый наркоти- ческий препарат-стимулятор, вызывая развитие не только привыкания, но и зависимости. Наркотическая зависи- мость — это ситуация, когда организм включает поступаю- щий извне препарат в свой метаболизм, т. е. начинает «рас- считывать» на его постоянный приток. При отказе от препара- та происходит сбой в использующих его системах мозга: наблюдается резкое ухудшение самочувствия, депрессия {абс- тинентный синдром или синдром отмены)- Человеку, по- павшему в зависимость, наркотик необходим уже не столько для того, чтобы почувствовать бодрость и эйфорию, сколько для возврата хотя бы к относительно нормальному уровню жизнедеятельности. Наиболее известным антагонистом никотиновых рецепто- ров является тубокурарин — активное действующее начало яда некоторых южноамериканских растений. Основным «мес- том приложения» его влияния являются нервно-мышечные синапсы (рис. 3.27, вариант /). При этом происходит последо- вательное расслабление и паралич мышц пальцев, затем глаз, рук и ног, шеи, спины и, наконец, дыхательных. Мускариновые рецепторы являются метаботропными (рис. 3.26, б); они связаны с G-белками, и присоединение к ним ацетилхолина приводит к синтезу вторичных посредников. Выделяют две основные локализации мускариновых ре- цепторов: синапсы, образуемые постганглионарными (в основ- ном парасимпатическими) вегетативными волокнами и ЦНС. В первом случае в качестве вторичных посредников использу- ются инозитолтрифосфат и диацилглицерол; во втором — цГМФ. Ионные последствия возбуждения мускариновых ре- цепторов весьма разнообразны: в сердце наблюдается увеличе- ние проводимости для ионов К+, что приводит к гиперполяри- зации и снижению частоты сокращений; в гладких мышцах отмечаются изменения проводимости как для К+, так и для Na+ (возможна гипер- или деполяризация в зависимости от конкретного органа). В ЦНС отмечается снижение проницаемости мембраны для К+ (деполяризация; возбуждающее действие), но синап- сы, содержащие мускариновые рецепторы, могут распола- гаться как на тормозных, так и на возбуждающих нейронах коры и базальных ганглиев. В связи с этим последствия бло- кады либо активации мускариновых рецепторов на поведен- ческом уровне оказываются очень индивидуальны; их выра- женность и направленность зависит от конкретной химиче- ской структуры того или иного препарата. Эффекты мускарина носят преимущественно парасимпа- тический характер: при отравлении мухоморами наблюдается тошнота, повышенное пото- и слюноотделение, слезотечение, боли в животе, снижение артериального давления и сердечной активности. Классическим антагонистом мускариновых рецепторов яв- ляется атропин — токсин белены и дурмана. Его перифериче- ские эффекты прямо противоположны действию мускарина: происходит снижение тонуса мышц желудочно-кишечного тракта, учащается сердцебиение, прекращается слюноотделе- ние (сухость во рту), расширяются зрачки, наблюдаются и центральные эффекты (двигательное и речевое возбуждение, галлюцинации).
Инактивация ацетилхолина происходит непосредственно в синаптической щели. Ее осуществляет фермент ацетилхоли- нэстераза, разлагающий медиатор до холина и остатка уксус- ной кислоты, затем холин всасывается в пресинаптическое окончание и может вновь использоваться для синтеза ацетил- холина. Ацетилхолинэстераза имеет активный центр, узнающий холин, и один активный центр, «отрывающий» ацетильную группу от исходной молекулы. Последний часто является мес- том атаки специфических блокаторов (рис. 3.27, вариант //). Примером подобного блокатора служит прозерин (неостиг- мин), применяемый при миастении, которая встречается при- мерно у трех человек на тысячу (чаще у женщин). Симптома- ми заболевания служат быстрая мышечная утомляемость, не- произвольное опускание век, замедленное жевание. Такие больные очень чувствительны к тубокурарину, а введение бло- каторов ацетилхолинэстеразы ослабляет патологические про- явления. В настоящее время известно, что у значительной части больных миастенией число никотиновых рецепторов примерно на 70% меньше, чем в норме. Причина этого состо- ит в том, что иммунная система больного вырабатывает анти- тела на никотиновые рецепторы. Эти антитела ускоряют раз- рушение рецепторов на мембране, ослабляя передачу в нерв- но-мышечном синапсе (рис. 3.27, вариант IV). Прозерин и сходные с ним препараты называют обратимы- ми блокаторами ацетилхолинэстеразы, их действие прекра- щается через несколько часов после введения, кроме того, су- ществуют необратимые блокаторы того же фермента. В этом случае вещество, нарушающее работу ацетилхолинэстеразы, вступает с белком в устойчивую химическую связь и выводит его из строя. Таким образом действуют фосфорорганические соединения, применяемые как препараты против насекомых (инсектициды): хлорофос, тиофос и сходные с ними соедине- ния могут вызвать у человека сужение зрачков, потливость, снижение артериального давления, подергивания мышц. Еще более сильными агентами-блокаторами являются раз- личные нервно-паралитические газы (зарин): легко проникая через все барьеры организма, они вызывают судороги, потерю сознания и паралич. Смерть наступает от остановки дыхания. Для немедленного ослабления эффектов отравляющих газов рекомендуется использование атропина; для восстановления деятельности ацетилхолинэстеразы — особые вещества-реак- тиваторы, «отрывающие» блокатор от фермента. Другим примером разрушительного действия на ацетилхо- линергический (использующий ацетилхолин в качестве меди- атора) синапс являются нейротоксины змей. Например, яд коб- ры содержит альфа-нейротоксин, необратимо связывающийся с никотиновым рецептором и блокирующий его, а также бе- та-нейротоксин, который тормозит выделение медиатора из пресинаптического окончания (рис. 3.27, варианты /, III).