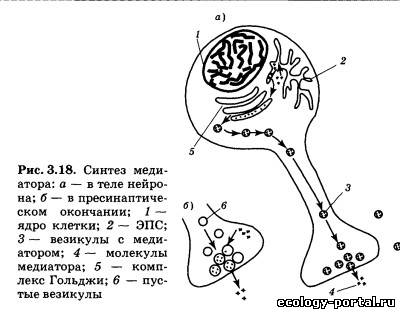
Жизненный цикл медиаторов нервной системы включает следующие «стандарные» стадии: синтез, загрузку в везику- лы и транспорт в пресинаптическое окончание; выделение в синаптическую щель; связывание с рецептором на постсинап- тической мембране; инактивацию. Образование медиатора часто происходит непосредственно в пресинаптическом окончании. Это возможно тогда, когда про- цесс синтеза является химически относительно простым (осу- ществляется в 1—2 стадии) и не требует каких-либо труднодос- тупных предшественников. Если эти условия не выполняют- ся, образование медиатора идет в теле нейрона (рис. 3.18). Это наиболее характерно для пептидных медиаторов, возникаю- щих в результате «вырезания» из более крупных белковых молекул. С синтезом каждого конкретного медиатора связаны спе- цифические ферменты, осуществляющие соответствующие реакции. От их количества и активности в конечном счете за- висит активность медиаторной системы. Другой важный фак- тор — наличие молекул-предшественниц. В этом случае дефи- цит медиаторов, образуемых из незаменимых веществ (тех, которые можно получить только с едой), может иметь пище- вое происхождение. Синтезированные в теле нейрона молекулы медиатора пе- реносятся сначала в ЭПС, а затем в комплекс Гольджи, кото- рый обеспечивает экзоцитоз медиаторов, предварительно упа- ковывая их в мембранные пузырьки-везикулы. Размер пу- зырьков и количество в них молекул медиатора стабильны в каждом конкретном нейроне.
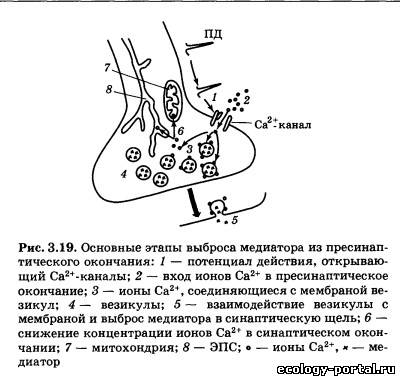
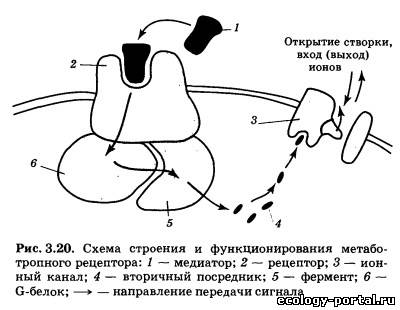
Образовавшиеся везикулы переносятся в пресинаптиче- ские окончания. Ключевую роль в этом процессе играют на- правляющие микротрубочки. Пузырьки с медиатором дви- жутся по этим «рельсам» с помощью механизмов, сходных с работой сократимых мышечных белков. Скорость такого транспорта довольно велика — до нескольких см/ч. В случае, когда медиатор синтезируется сразу в пресинапти- ческом окончании, комплекс Гольджи способен формировать пустые везикулы, которые аналогичным образом переносятся по аксону. Заполнение таких пузырьков медиатором осуществ- ляется непосредственно в пресинаптическом окончании (за счет работы специальных молекулярных насосов (рис. 3.18, б)). Везикулы — это не только удобная форма транспорта ве- ществ, но и способ упорядочить, сделать количественно ста- бильным выброс медиатора в синаптическую щель. Число скапливающихся в пресинаптическом окончании везикул из- меряется десятками тысяч, что также стабилизирует процес- сы передачи сигналов; истощение запасов медиатора даже при интенсивном проведении информации происходит весьма ред- ко (обычно на фоне действия специальных фармакологиче- ских агентов). Каждый нейрон производит только один основной меди- атор (ацетилхолин, дофамин и т. п.). Однако нередко можно обнаружить присутствие в пресинаптическом окончании и других веществ, способных к передаче нервных сигналов. Это комедиаторы (например, пептиды); они обнаруживаются в очень небольших количествах и обычно находятся в везику- лах, отличающихся по форме и размеру от пузырьков с основ- ным медиатором. Выброс содержимого везикул запускается в момент прихо- да в пресинаптическое окончание потенциала действия и ре- ализуется в несколько этапов. Первый из них заключается в от- крытии потенциал-зависимых Са2+-каналов, которые располо- жены на «внешней» мембране пресинаптического окончания и открываются при его деполяризации в момент прихода ПД (рис. 3.19). В результате наблюдается вход в клетку определен- ной порции ионов Са2+, и их содержание внутри окончания воз- растает в 10—100 раз. Чем выше концентрация Са2+ во внеш- ней среде, тем больше число вошедших ионов; через эти же ка- налы способны проникать ионы Mg2+, конкурируя с кальцием. Следовательно, появление в межклеточной среде магния умень- шает итоговое число вошедшего в окончание кальция. Основное назначение ионов Са2+ в пресинаптическом окон- чании — это воздействие на сложный белковый комплекс, встроенный в мембрану везикул. Этот комплекс включает бел- ки, ответственные за фиксацию пузырька в цитоплазме и за его контакт с пресинаптической мембраной. Под действием Са2+ (для этого нужно четыре иона) везикула приходит в дви- жение. Достигая пресинаптической мембраны, пузырек «сли- пается» с ней, в результате чего медиатор попадает в синапти- ческую щель. Весь этот процесс протекает очень быстро — в течение 1—5 мс, а примерно через 10 с можно наблюдать процесс восстановления везикул: они отделяются от мембраны и возвращаются в пресинаптическое окончание. В дальнейшем эти пустые пузырьки могут быть вновь заполнены медиатором. Запустить выброс содержимого везикул чрезвычайно важ- но, но не менее важно быстро остановить этот процесс. Только в этом случае возможно точное соответствие между числом пришедших в пресинаптическое окончание ПД и количест- вом выделившегося медиатора. Функцию остановки выброса выполняют особые молекулярные насосы, удаляющие ионы Са2+ из цитоплазмы окончания. Такие насосы находятся на мембранах каналов ЭПС и митохондрий. Перенося кальций внутрь этих органоидов, они прекращают его действие на ве- зикулы. Отравление Са2+-насосов ведет к гиперактивности синапса, продолжающейся до полного истощения запасов медиатора. Аналогичное действие оказывают токсины, блокирующие по- тенциал-зависимые Са2+-каналы в открытом положении. Токсин ботулиновой бактерии (ботулотоксин) известен как соединение, вызывающее тяжелейшие пищевые отравления. Проникая в синапс, он блокирует белки, отвечающие за кон- такт везикулы с пресинаптической мембраной, в результате чего прекращается всякая передача нервного сигнала, разви- ваются параличи. Попав в синаптическую щель, медиатор менее чем за 1 мс вступает во взаимодействие с пресинаптической мембраной, соединяясь с встроенными в нее специализированными белко- выми рецепторами. Пространственная организация рецепто- ра предусматривает существование у него активного центра — углубления в белковом клубке, имеющего определенную форму и распределение зарядов. Ему строго соответствует простран- ственная конфигурация медиатора и распределение зарядов на его молекуле. В результате активный центр рецептора и медиатор способны формировать комплекс. Непосредствен- ным следствием этого является возбуждение рецептора, а за- тем — развитие постсинаптических потенциалов и запуск ПД. Выделяют два типа рецепторов — ионотропные и метабо- тропные. Возбуждение метаботропного рецептора выражается в изменении внутриклеточного метаболизма, т. е. течения био- химических реакций. С внутренней стороны мембраны к та- кому рецептору присоединен целый ряд других белков, вы- полняющих ферментативные и частью передающие («посред- нические») функции (рис. 3.20). Белки-посредники относятся к G-белкам. Под влиянием возбужденного рецептора G-белок воздействует на белок-фермент, обычно переводя его в «рабо- чее» состояние. В результате запускается химическая реак- ция: молекула-предшественник превращается в сигнальную молекулу — вторичный посредник.
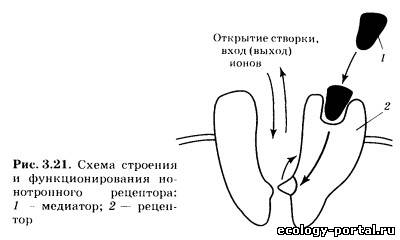
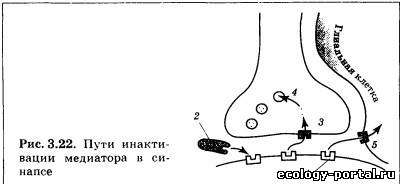
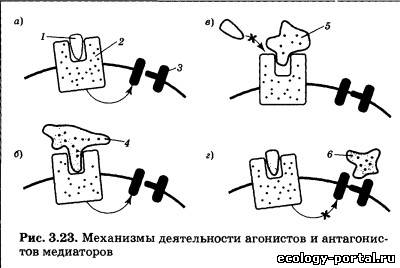
Вторичные посредники — это мелкие, способные к быстро- му перемещению молекулы или ионы, передающие сигнал внутри клетки. Этим они отличаются от «первичных посред- ников» — медиаторов и гормонов, передающих информацию от клетки к клетке. Наиболее известным вторичным посредни- ком является цАМФ (циклическая аденозинмонофосфорная кислота), образуемая из АТФ с помощью фермента аденилат- циклазы. Похожа на него цГМФ (гуанозинмонофосфорная кислота). Другими важнейшими вторичными посредниками являются инозитолтрифосфат и диацилглицерол, образуе- мые из компонентов клеточной мембраны под действием фер- мента фосфолипазы С. Чрезвычайно велика роль Са2+, входя- щего в клетку снаружи через ионные каналы или высвобож- дающегося из особых мест хранения внутри клетки («депо» кальция). В последнее время много внимания уделяется вто- ричному посреднику N0 (оксиду азота), который способен пе- редавать сигнал не только внутри клетки, но и между клетка- ми, легко преодолевая мембрану, в том числе от постсинапти- ческого нейрона к пресинаптическому. Заключительный шаг в проведении химического сигнала — воздействие вторичного посредника на хемочувствительный ионный канал. Это воздействие протекает либо непосредствен- но, либо через дополнительные промежуточные звенья (фер- менты). В любом случае происходит открытие ионного канала и развитие ВПСП либо ТПСП. Продолжительность и амплиту- да их первой фазы будет определяться количеством вторично- го посредника, которое зависит от количества выделившегося медиатора и длительности его взаимодействия с рецептором. Таким образом, механизм передачи нервного стимула, ис- пользуемый метаботропными рецепторами, включает в себя несколько последовательных этапов. На каждом из них воз- можна регуляция (ослабление либо усиление) сигнала, что де- лает реакцию постсинаптическои клетки более гибкой и адап- тированной к текущим условиям. Вместе с тем это же при- водит к замедлению процесса передачи информации. Вот почему в ходе эволюции возникла потребность в более быст- ром пути проведения сигналов, в результате чего появились ионотропные рецепторы. В случае ионотропного рецептора чувствительная молекула содержит не только активный центр для связывания медиато- ра, но также ионный канал (рис. 3.21). Воздействие «первич- ного посредника» на рецептор приводит к быстрому открыва- нию канала и развитию постсинаптического потенциала. Инактивация — заключительный этап жизненного цикла медиатора. Смысл этой стадии состоит в прекращении его дей- ствия на рецептор (прерывание передачи сигнала). Процессы инактивации медиатора реализуются при участии специали- зированных ферментов и транспортных белков (рис. 3.22). В более простом случае инактивация осуществляется пря- мо в синаптической щели, когда фермент 2 быстро разрушает все свободно «плавающие» молекулы медиатора. Кроме этого, медиатор может быть удален с активных центров постсинап- тических рецепторов 1 еще двумя способами: путем обратного всасывания в пресинаптическое окончание, которое осуществ- ляется особыми белками-насосами 3 и путем всасывания в глиальные клетки, которое также происходит за счет деятель- ности белков-насосов 5. В случае переноса внутрь глиальных клеток медиатор раз- рушается специализированным ферментом. В случае возврата в пресинаптическое окончание {«обрат- ный захват») он также может быть разрушен, но может и по- вторно «загружаться» в пустые везикулы 4. Это позволяет наиболее экономно расходовать те медиаторы, синтез которых связан с определенными проблемами (недостаток предшест- венника, длинная цепочка реакций). Скорость процесса инактивации определяет общее время воздействия медиатора на рецептор, от которого в конечном итоге зависит амплитуда постсинаптических потенциалов, за- пуск ПД и продолжение проведения сигнала по нейронной се- ти. При повреждении элементов системы инактивации наблю- дается значительное увеличение эффективности синаптиче- ской передачи, так как выделившийся медиатор существенно дольше воздействует на рецепторы и амплитуда ВПСП либо ТПСП заметно возрастает. Вещества, влияющие на различные этапы жизненного цикла медиаторов, имеют огромное значение для жизни че- ловека. Именно они образуют группу психотропных препа- ратов — соединений, влияющих на различные аспекты деятельности мозга: общий уровень активности, память, эмо- циональные переживания. При этом наиболее часто использу- ются вещества, изменяющие взаимодействие рецептора и ме- диатора, а также влияющие на хемочувствительные ионные каналы. При введении молекул, сходных по структуре с медиато- ром, наблюдается их соединение с активными центрами соот- ветствующих рецепторов и последующее возбуждение рецеп- торов (рис. 3.23). Медиатор 1 присоединяется к рецептору 2, что приводит к воздействию на ионный канал 3 (направление воздействия отмечено стрелкой; рис. 3.23, а); агонист 4 присо- единяется к рецептору, что также приводит к передаче сигна- ла на ионный канал (рис. 3.23, б); конкурентный антагонист 5 не позволяет медиатору соединиться с рецептором (отмечено перечеркнутой стрелкой; рис. 3.23, в); неконкурентный анта- гонист 6 блокирует ионный канал, что также не позволяет развиться эффектам медиатора (отмечено перечеркнутой стрелкой; рис. 3.23, г). В результате эффект применяемого препарата оказывается аналогичен действию самого медиато- ра. Вещества такого рода называют агонистами медиатора, их влияние на синапс нередко оказывается очень длительным и эффективным. Это объясняется тем, что прочность связыва- ния агониста с рецепторами нередко больше, чем у медиатора, а системы инактивации не способны быстро опознать агонист и убрать его из синаптической щели. В более сложном случае вводимые молекулы лишь частич- но похожи на медиатор. Тогда, соединяясь с активными цент- рами рецепторов, они будут их занимать (прекращать к ним доступ медиатора; конкурировать с ним), но не будут возбуж- дать рецептор. В результате эффект применяемого препарата будет противоположен действию медиатора. Вещества такого рода называют конкурентными антагонистами медиатора. Существует также понятие неконкурентного антагониста (вводимый препарат мешает работе медиатора, блокируя хе- мочувствительные ионные каналы). Часть агонистов и антагонистов медиаторов являются ве- ществами природного происхождения. Их существование — результат длительных эволюционных процессов, в ходе кото- рых одни живые организмы (особенно растения) «изобретали» вещества, защищающие их от поедания другими организма- ми. Природными психотропными препаратами являются так- же яды животных-охотников (змеи, пауки). Вторая часть агонистов и антагонистов — синтетические соединения, создаваемые человеком. В ходе их разработки хи- микам и фармакологам приходится учитывать целый ряд тре- бований. Во-первых, в структуре такого вещества должен при- сутствовать «ключевой» участок, соответствующий молекуле медиатора. Во-вторых, такой препарат должен быть устойчив к действию систем инактивации. В-третьих, он должен прони- кать через барьеры организма — гематоэнцефалический и же- лательно кишечный. Только в этом случае можно достичь мозга при системном введении — в виде таблетки либо инъек- ции. В настоящее время агонисты и антагонисты медиаторов (а также соединения, влияющие на синаптическую передачу другими путями) широко применяются в клинике. Вместе с тем в больших дозах многие из них являются наркотиками и ядами, что также свидетельствует о необходимости их серьез- ного изучения.