 |
Экологический портал Главная | Регистрация | Вход |
Суббота, 18.05.2024, 02:26 Приветствую Вас Гость |
|
|
|
|||||||||||||||||||||||||||||||||||||||






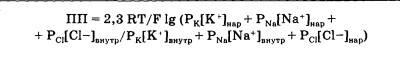
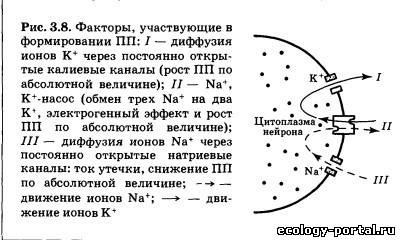



 |
Copyright ecology-portal.ru © 2024 |  |
|
Экологический портал Главная | Регистрация | Вход |
Суббота, 18.05.2024, 02:26 Приветствую Вас Гость |
|
|
|
|||||||||||||||||||||||||||||||||||||||
|
Copyright ecology-portal.ru © 2024 | |